{kind=link}
[pdf] Marine Biology (2006) 149:689-701
[url] Related article in Limnology & Oceanography
Introduction
The Santa Barbara Channel Islands (Santa Cruz Island and nearby Santa Rosa Island)
are located in an extremely dynamic oceanographic setting.
Strong gradients in environmetal conditions are observed over relatively small spatial
scales (i.e. 100 km). These gradients in environmental conditions are stable
over time with the south shores of Santa Cruz Island (SCI) experiencing strong wave
action and the influx of warmer subtropical waters.
The north shores of SCI are relatively protected for wave action and experience
colder water temperatures originating in the Point Conception upwelling center.
The patchy environmetal setting creates a unique opportunity to test current theories
of community regulation in rocky intertidal communities.
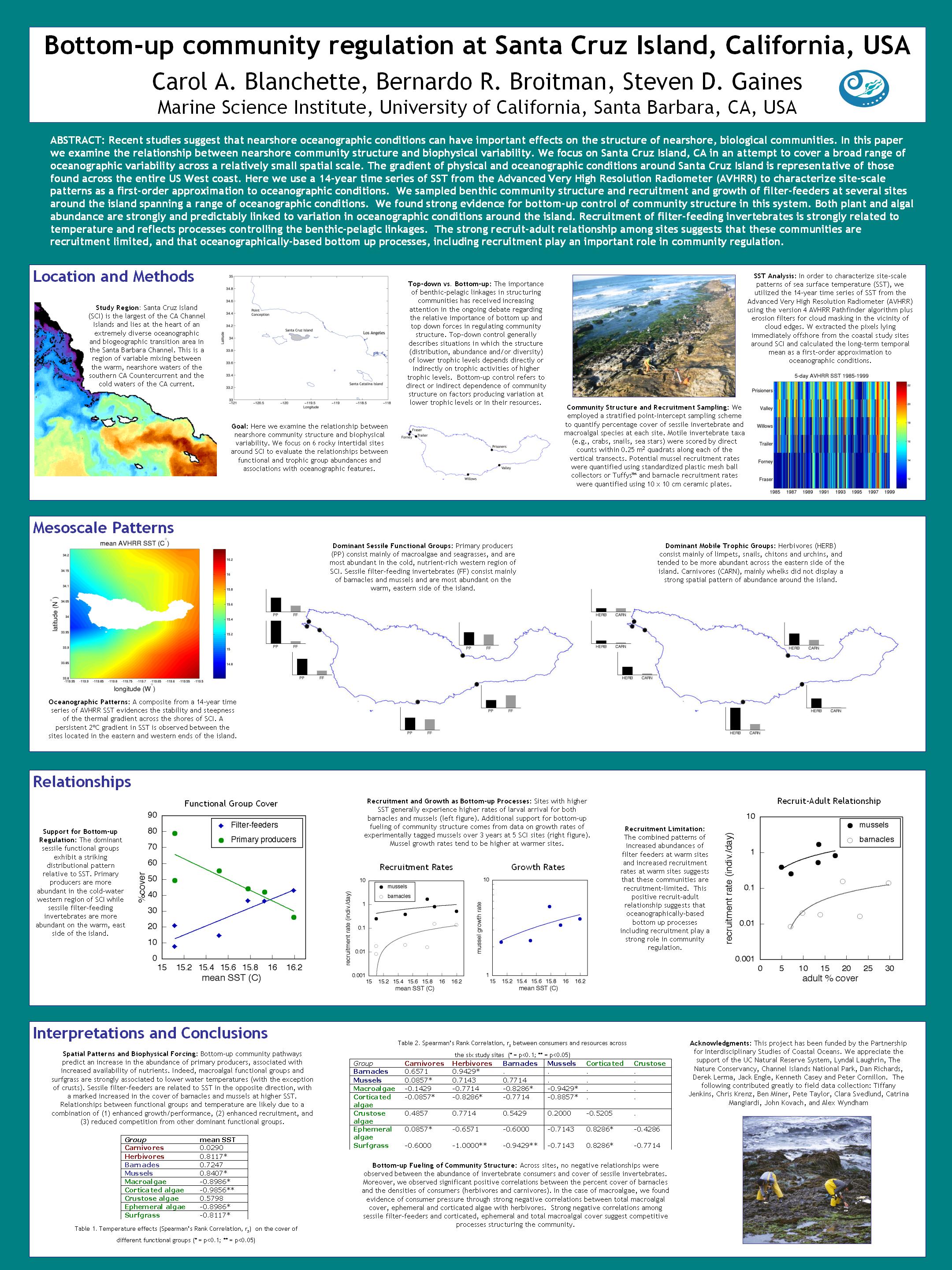
A composite from a 6-year time series of sea surface temperature (SST) from the Advanced
Very High Resolution Radiometer (AVHRR) satellite sensor evidences the stability of the
thermal gradientacross the shores of SCI (fig. 1).
|
| Figure 1. Mean SST from the 6-year AVHRR time series of weekly means. The nominal pixel resolution is 1 km |
Methods
SST analyses
In order to characterize site-scale patterns of SST, we utilized the 6-year time series
of SST from the Advanced Very High Resolution Radiometer (AVHRR)
(Kenneth S. Casey and Peter Cornillon. A comparison of in situ-based sea surface temperature
climatologies. Journal of Climate, Vol 12, No 6, pp. 1848-1863, 1999).
Correlation analyses
We studied the role of different processes structuring the intertidal community through
Spearman rank correlation (rs) analyses (Zar, 1996).
The influence of biophysical variability was examined through the correlation between the
long-term mean SST and the abundance of different functional groups across all sites.
Results
Spatial community patterns
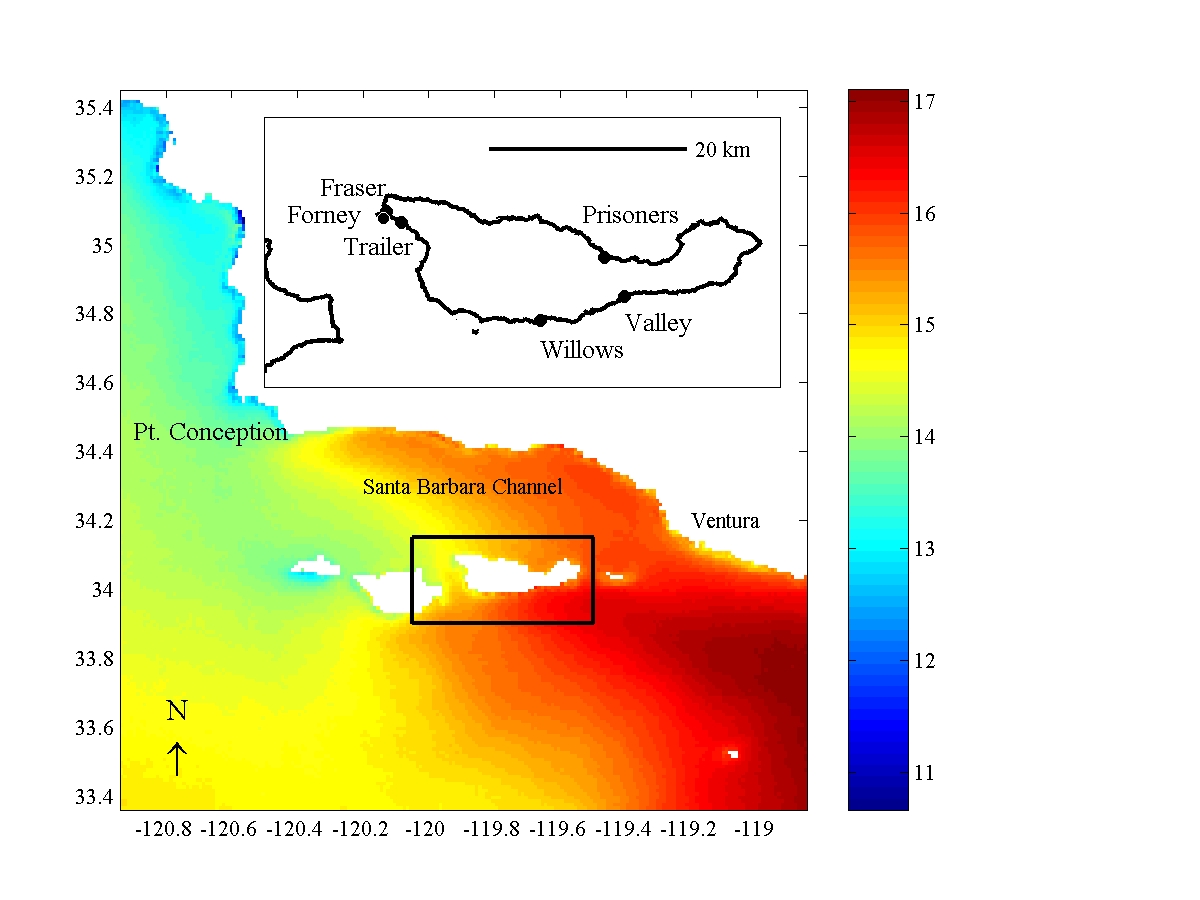
Patterns if abundance of functional groups display a marked spatial pattern. Primary producers are
more abundant in the western part of SCI while sessile filter-feeding invertebrates are more abundant
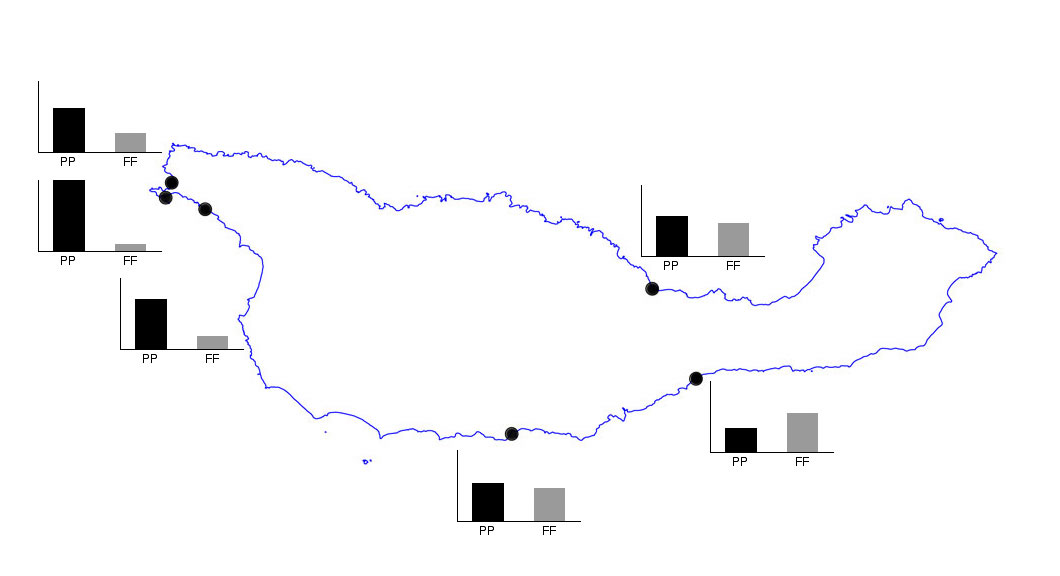
on the east side of the island (fig. 2). A similar pattern is observed in the abundance of
 |
 |
Figure 2.Spatial pattern of abundance of primary space occupiers. | Figure 3.Spatial patterns of abundance of invertebrate consumers. |
| Group | mean AVHRR SST |
| Seastars | 0.9959** |
| Herbivores | 0.8117* |
| Barnacles | 0.7247 |
| Mussels | 0.8407* |
| Macroalgae | -0.8986* |
| Corticated algae | -0.9856** |
| Crustose algae | 0.5798 |
| Ephemeral algae | -0.8986* |
| Surfgrass | -0.8117* |
Sites with higher sea surface temperature (SST) exhibit higher rates of larval arrival (fig. 4). The relationship with SST is extremely high for both mussel and barnacle larvae. Adult abundances follow an equally consistent pattern with SST (Table 2). The adult abundance pattern that may be more related to recruitment limitation, as evidenced by the strong adult-recruit relationship (fig. 5).
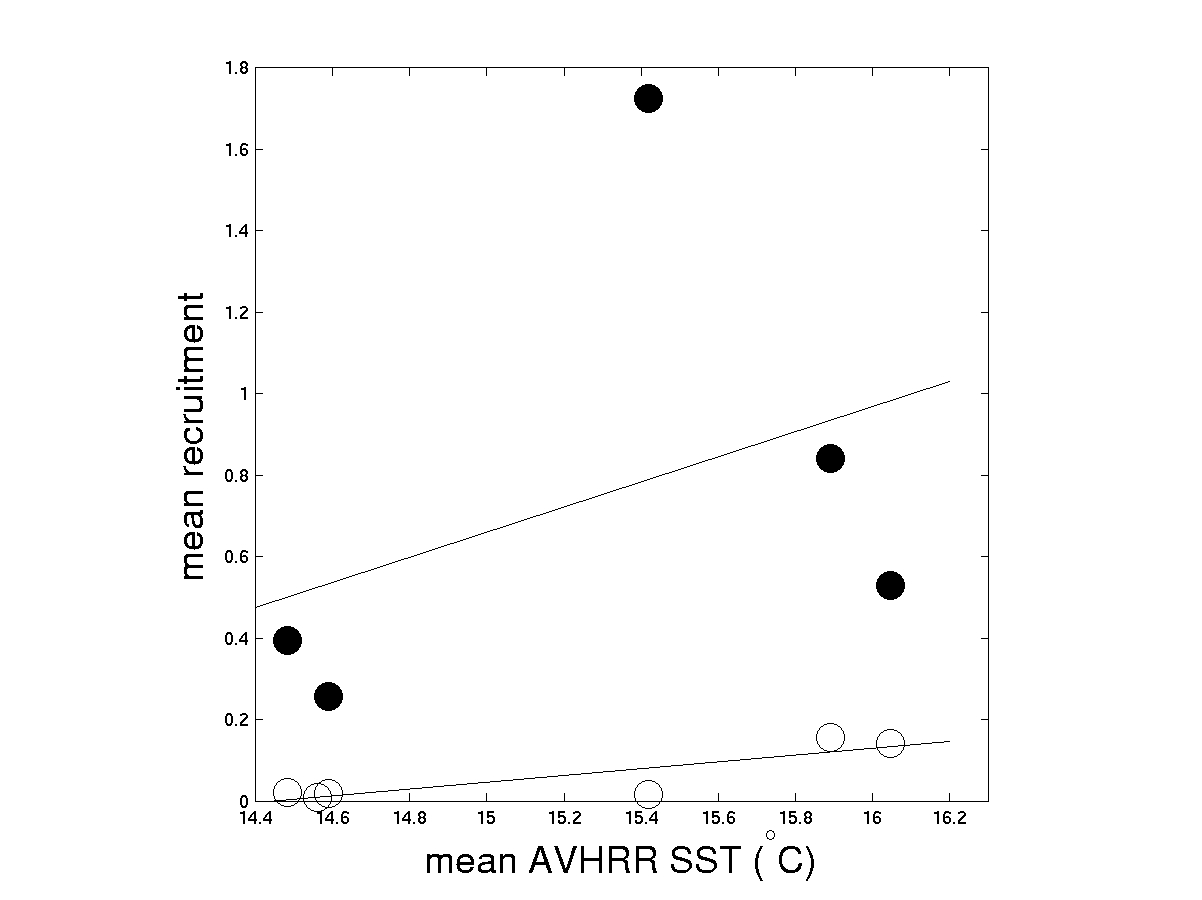 |
| Figure 4. SST vs. larval arrival. Barnacles (Chthamalus spp. ,open circles) and mussels (Mytilus spp., black circles). |
Additional support for a "bottom-up fueling" of community structure comes from growth rate data obtained at five sites in SCI. It can be seen that a strong relationship is observed with growth rate averaged across all years (fig. 5).
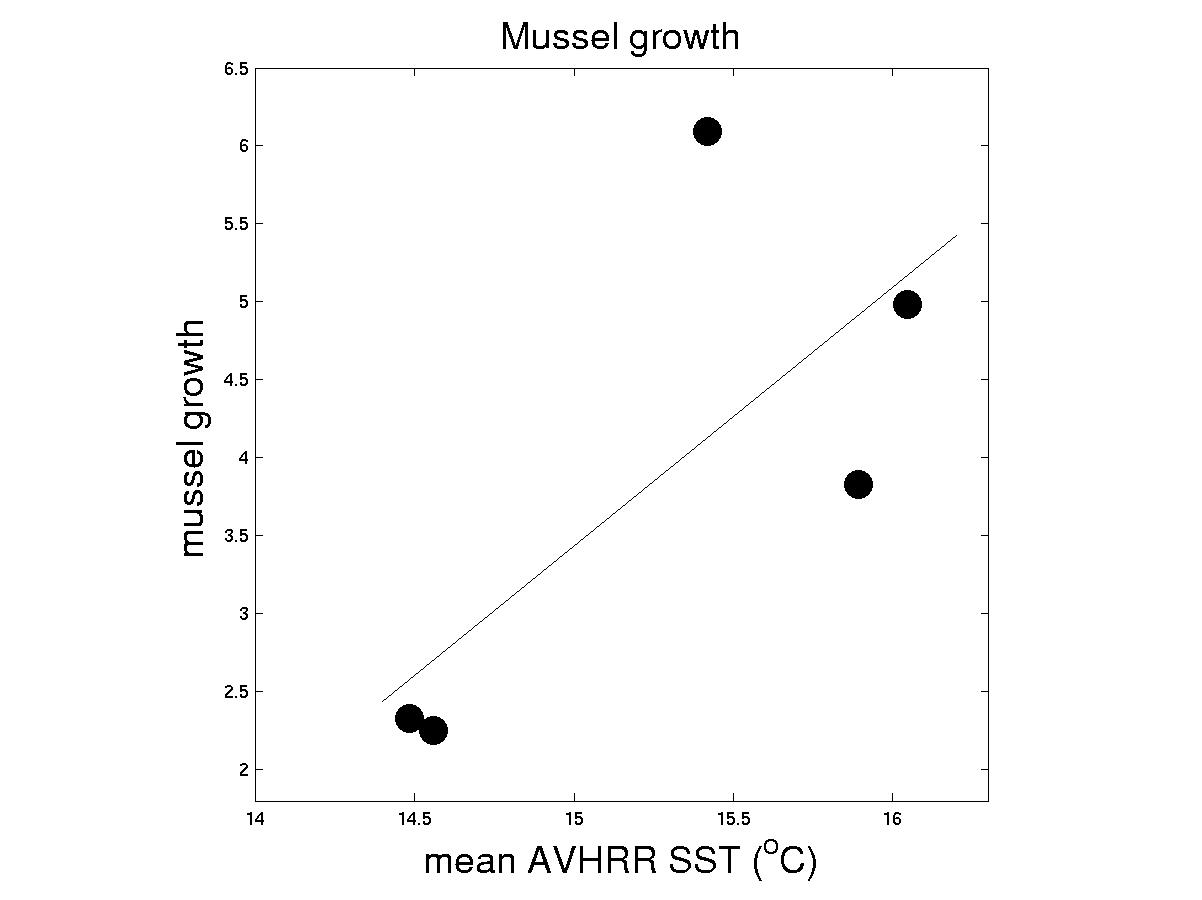 |
| Figure 5. Temperature effects on the multi-year mean of growth of transplanted mussels. r = 0.6150 |
Community patterns
Across sites, no negative relationships were observed between the abundance of invertebrate consumers and
cover of sessile invertebrates. Moreover, we observed significant positive correlations between the
percent cover of barnacles and the densities of consumers (herbivores and carnivores).
In the case of macroalgae, we found evidence of consumer pressure through strong negative correlations
between total macroalgal cover, ephemeral and corticated algae with herbivores.
Strong negative correlations among sessile filter-feeders and corticated,ephemeral and total macroalgae cover
suggest competitive processes structuring the community. Some interesting relationships are observed
which are in wide agreement with the patterns reported for Chile. Corticated algae have strong negative
correlations with sessile invertebrates (mussels and barnacles). Mussels and barnacle adults are not
negatively, but positively correlated across sites.
| Group | Carnivores | Herbivores | Barnacles | Mussels | Corticated | Crustose |
| Barnacles | 0.6571 | 0.9429* | . | . | . | . |
| Mussels | 0.0857* | 0.7143 | 0.7714 | . | . | . |
| Macroalgae | -0.1429 | -0.7714 | -0.8286* | -0.9429* | . | . |
| Corticated algae | -0.0857* | -0.8286* | -0.7714 | -0.8857* | . | . |
| Crustose algae | 0.4857 | 0.7714 | 0.5429 | 0.2000 | -0.5205 | . |
| Ephemeral algae | 0.0857* | -0.6571 | -0.6000 | -0.7143 | 0.8286* | -0.4286 |
| Surfgrass | -0.6000 | -1.0000** | -0.9429** | -0.7143 | 0.8286* | -0.7714 |